Chu kỳ tế bào và cơ chế sửa sai trong sao chép: qúa trình bệnh lý, cơ chế bệnh sinh, hoạt động cơ quan tổ chức
Tin liên quan
- 10 hoạt động thú vị không nên bỏ lỡ khi tới Hội An: Số 8 nhiều người Việt còn chưa biết (30/07/2022)
- 15 bức ảnh kỳ quặc, nhìn mãi mới hiểu sự thật (11/09/2022)
- Cần quản lý chặt chẽ hoạt động nuôi cá bè trên hồ Trị An (22/07/2022)
- Di dời lò mổ sau 26 năm hoạt động (27/03/2023)
- Phát hiện ra cơ quan quan trọng trong bộ não người mà chúng ta chưa hề biết trước đây (09/01/2023)
Chu kỳ tế bào
Một chu kỳ sinh học tế bào tức là giai đoạn giữa hai lần phân chia gồm 4 pha: M, G1, S, G2. Chu kỳ phân chia kéo dài khoảng 16-24 giờ tuỳ theo mỗi loại tế bào.
M (mitosis): Hoạt động phân chia tế bào hay nhân đôi tế bào
G1 (gap): Có sự tích luỹ vật chất nội bào và năng lượng, kết thúc ở điểm tới hạn R (restriction) vài giờ trước khi chuyển từ G1 sang S. Một khi tế bào đi qua được điểm R sẽ đi qua các pha khác để thực hiện được phân bào.
S (synthesis): Giai đoạn tổng hợp DNA, lượng DNA tăng gấp đôi (từ 23 đôi thành 46 đôi)
G2: Quy trình được hoàn tất và chuẩn bị sang pha sau
M (mitosis): Mỗi cặp kép nhiễm sắc thể chia đôi, đi về 2 cực tạo thành 2 tế bào con y hệt tế bào mẹ.
Sau khi phân đôi 2 tế bào con có thể tiếp tục chu trình ấy hoặc đi vào thời kỳ nghỉ là G0.
G0: Các thời gian dừng của chu kỳ tế bào là để sửa chữa DNA cho tế bào sống sót và không tiến triển thành ung thư: sự ngừng chu kỳ tế bào ở G1/S tránh được sự tái sao của các DNA thương tổn, sự dừng ở G2/M tránh được sự ngưng tập của các nhiễm sắc thể bị thương tổn.
Cơ chế sửa sai trong chu kỳ tế bào
Ngay từ những năm 1960 Leland Hartwell đã phân lập được nhiều loại tế bào có đột biến gen ở một loại nấm men (Saccharomyces cerevisae). Bằng cách tái hợp chúng với nhau đã phát hiện ra sự kiểm soát phân chia tế bào nằm trên NST do hằng trăm gen khác nhau. tên gọi chung là gen CDC (cell division cycle genes) với chữ số theo sau cho từng thứ. Trong số các gen bày có hai loại quan trọng nhất là điểm khởi phát chu kỳ và những điểm quan trọng mà khi bị hư hỏng thì gián đoạn phân chia tế bào, gọi là điểm kiểm soát (check point)
Sửa sai trong sao chép
Thực nghiệm dùng các nucleotid, DNA polymerase để tổng hợp DNA thì nhận thấy sai sót xảy ra rất cao (1x10-5) trong khi sao chép tự nhiên lại thấp hơn nhiều. DNA của E. coli có 3x106 cặp bazơ như vậy thì mỗi lần sao chép phải có 30 sai sót xảy ra dẫn đến sự đột biến nhưng điều này không xảy ra như vậy trong tự nhiên. Nguyên nhân chính của sự chính xác này là hiện tượng sửa chữa DNA xảy ra ở mọi tế bào bình thường trong cơ thể sinh vật bậc cao.
Theo dõi tần số đột biến ở các quần thể lớn cho thấy tỷ lệ đột biến chỉ ở 1x10-9, như vậy ở người mỗi lần sao chép chỉ có 3 sai sót xảy ra cho mỗi DNA, như vậy kết luận cơ thể sinh vật đã có những cơ chế sửa sai tế bào.
Hàng chục loại enzym khác nhau đã tham gia vào quá trình sửa chữa các DNA tổn thương. Chúng nhận biết chọn lọc một bazơ bị thay đổi, loại bỏ nucleotid mang nó bằng cách cắt ra khỏi chuỗi DNA, sau đó thay bằng một nucleotid mang bazơ chính xác bổ sung và gắn DNA lại. Người ta cho rằng với cơ chế này cho phép sửa chữa 99,9% các sai sót.
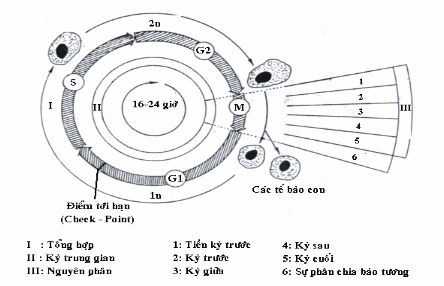
Hình: Chu kỳ tế bào.
Mở đầu khi tế bào chuyển sang pha G1, Harwell phát hiện là nó sẽ bị ngăn lại khi thiếu chất dinh dưỡng hoặc chất chuyển hoá trung gian, có sự tham gia của gen RNA polymerase (primase) tạo ra đoạn mồi đầu tiên để cho DNA polymerase tiếp tục quá trình tái sao DNA cũng như loại bỏ mẩu mồi ấy. Khi tế bào qua khỏi điểm này (điểm khởi phát, điểm R: restriction) thì sẽ không quay trở lại được và phải đi tiếp sang pha S. Những rối loạn xảy ra ở các bước sau tế bào sẽ bị loại bỏ bằng cơ chế ch*t theo chương trình (apoptosis).
Các sự kiện của chu kỳ tế bào diễn tiến theo một trật tự nhất định, sự kiện trước phải được hoàn tất tốt đẹp thì sự kiện sau mới tiếp tục xảy ra. Các cơ chế kiểm soát nhờ hoạt động của những gen nằm ở những nơi gọi là điểm kiểm soát. Khi các điểm kiểm soát bị loại bỏ sẽ gây ch*t tế bào, sai lệch trong phân bố nhiễm sắc thể hay các phần tử tế bào hoặc tăng nhạy cảm với các yếu tố môi trường.
Sau này Harwell đã tìm ra được nhiều gen của điểm kiểm soát. Ví dụ như tế bào nấm có gen RAD53 khi bị chiếu tia thì sẽ gây ngừng ở G2 cho đến khi tổn thương được sửa chữa xong. Ở người có gen p53 trên nhiễm sắc thể 17 có vai trò đièu hoà tương tự như RAD53. Khi gen này do đột biến không hoạt động được thì tế bào dù bị hư hại (đột biến hay thay đổi do yếu tố sinh ung thư) vẫn cứ thế mà tiếp tục phân chia thành những thế hệ tế bào con có rối loạn y hệt gọi là ác tính.
Năm 1970 nhờ các kỹ thuật gen và sinh học phân tử đã phát hiện ra khi vượt qua điểm khởi động thì tế bào cần hoạt động của 2 gen cdc2 /cdc28 để bắt đầu tái sao DNA. Các gen này mã cho một protein kinase p34 là yếu tố điều hoà chủ yếu của phức hợp. Sau này người ta mới phát hiện ra vai trò của cyclin với p34cdc2 protein kinase
Năm 1980 R. Timothy Hunt đã phát hiện ra phân tử cyclin đầu tiên trong quá trình phân bào của con nhím biển, ức chế sự tổng hợp các protein này thì các giai đoạn phân bào không thực hiện được. Protein này hình thành rồi phân huỷ ngay trong mỗi chu kỳ tế bào nên mới có tên là cyclin (cycle). Hiện nay, cyclin tạo thành một họ gồm nhiều protein gần giống nhau có đặc tính chung là liên kết với các dưới đơn vị protein kinase trong họ cdc để trở nên các phức hợp hoạt động, gọi là protein kinase phụ thuộc cyclin (cdk). Để cho 1 pha chu kỳ tế bào đi qua được 1pha khác thì cdk này phải kết hợp với 1 cyclin nhất định.
Ví dụ:
Cyclin B cdk1: giúp tế bào đi từ G2 sang M Cyclin E cdk2: G1→S
Cyclin D cdk4: G0 → G1
Các gãy đơn (single strand breaks) và gãy kép (double strand breaks) của DNA làm tăng tổng hợp p53, làm tế bào dừng lại ở pha G1do ức chế được phức hợp Cyclin E cdk2 nên tế bào không đi vào pha S. Nếu thiếu p53, tế bào dễ phát triển thành ung thư.
Sửa sai khi không sao chép
Có khoảng 50 enzym chuyên rà soát, phát hiện và sửa các sai sót trên phân tử DNA.
Sự ch*t tế bào
Phân bào làm tăng số lượng tế bào với bộ gen giống nhau. Song song với sự phân chia tế bào còn có những cơ chế kiểm soát sự ch*t của tế bào theo chu kỳ phát triển. Có thể gặp trong một số quá trình S*nh l* như thoái triển S*nh l* phụ thuộc nội tiết tố, tế bào lympho T gây độc (Tc) làm ch*t tế bào đích....Ngoài ra, không kể đến những trường hợp ch*t tế bào do thoái hoá, hoại tử, sự teo ..v.v.
ch*t theo chương trình của tế bào (apoptosis nghĩa đen là rơi khỏi)
Trên tiêu bản nhuộm hematoxylin eosin cho thấy có sự biến tính của protein, chromatin đậm đặc. Cuối cùng có hiện tượng vỡ nhân do DNA bị gãy, tế bào nhanh chóng co lại và tạo ra các chồi bào tương, đứt gãy thành các “thể apotosis” chứa chất hoà tan và cơ quan tử của tế bào; các thể này được thực bào sau đó. Cơ chế này vẫn chưa hoàn toàn rõ ràng tuy nhiên thường bắt đầu bằng các thương tổn DNA, đặc biệt là hiện tượng gãy DNA chuổi kép, tăng giải phóng topoisomerase (có trong tế bào bình thường hoặc ác tính) có tác dụng tách đôi sợi DNA xoắn kép khi sao chép, sửa chữa các thương tổn DNA. Khi thiếu enzym này thì nhiễm sắc thể không phân ly và tế bào không phân bào được. Nhưng khi quá nhiều topoisomerase thì tế bào cũng sẽ ch*t theo chương trình khi có sự kết hợp giữa DNA chuổi kép và topoisomerase II.
Tóm lại, ở sinh vật đa bào sự kiểm soát phân bào và ch*t theo chương trình được kiểm soát bằng nhiều cơ chế phức tạp. Khi cơ chế kiểm soát này bị hỏng thì dẫn đến nguyên phân không giới hạn và tế bào hầu như không biệt hoá, xảy ra ở ung thư.
Sự kiểm soát di truyền đối với apoptosis
Bộ gen tuyến trùng Caenorhabditis elegans có 1031 gen, trong đó có 131 gen có phân định sẳn là ch*t theo chương trình. Có 14 gen khác nhau tham gia vào kiểm soát di truyền đối với apotosis.
Nếu có sự đột biến làm bất hoạt các gen ức chế sinh ung thư (ced -3 và ced -4) thì các tế bào trên sống sót tuy nhiên nếu đột biến gen kích thích sinh ung thư (ced -9) làm cho một số tế bào bình thường sống sót nay bị ch*t. Nghiên cứu cấu trúc chức năng của một protein của gen ced-9 thì thấy giống như sản phẩm của một proto-oncogen có tên là bcl-2. Ngoài ra p53 là sản phẩm của gen ức chế ung thư, là chất hoạt hoá một trong những con đường dẫn đến apoptosis và làm giảm protein bcl-2.
Apoptosis và bệnh lý khác
Sự ch*t theo chương trình của tế bào giữ vai trò cơ bản trong nhiều bệnh: điều hoà trực tiếp sự phát triển của khối u, góp phần ngăn chận hay làm chậm đi sự phát triển của bệnh AIDS. Nhiều bằng chứng cho thấy vai trò của apoptosis trong tuổi thọ, bệnh Alzheimer,.vv. Tuy được phát hiện không lâu nhưng apoptosis đã thu hút những nghiên cứu về điều trị ung thư và những bệnh lý khác.
Ung thư
(xem bài ung thư).
Tin liên quan
- Điều hòa kêu to ù ù khi hoạt động gây khó chịu: Đừng vội gọi thợ, làm thế này sẽ hết ngay (07/07/2022)
- Một xã, hơn 20 xưởng tái chế bao bì hoạt động trái phép (21/05/2022)
- Suy đa tạng vì 1 vết cắn từ loài vật hoạt động mạnh vào mùa hè: Người dân biết mà không sợ (22/06/2022)
- Tuần Lễ Cảm Ơn của UNIQLO diễn ra từ 10-16/6 với loạt hoạt động ý nghĩa (04/06/2022)
- UNIQLO đẩy mạnh hoạt động RE.UNIQLO nhân “Tuần Lễ Cảm Ơn” từ 10-16/6 (12/06/2022)
Tin cùng chuyên mục
Tin cùng nội dung
-
Nên ăn trứng luộc lòng đào thay vì rán, bởi trứng lòng đào không mất các dinh dưỡng thiết yếu cần thiết.
-
Lứa tuổi dậy thì được coi là cơ hội cuối cùng để các bậc cha mẹ có thể giúp con mình có chiều cao lý tưởng cũng như hoàn tất quá trình phát triển các bộ phận trên cơ thể thông qua con đường dinh dưỡng.
-
Thừa cân hoặc béo phì có liên quan tới tăng nguy cơ bị ung thư vú, ung thư đại tràng, thực quản, thận và có thể là các ung thư khác.
-
Dưới đây là 12 hoạt động thú vị giúp trẻ có một kỳ nghỉ hè giải trí nhưng vẫn đầy tính giáo dục.
-
Trong một cuộc thăm dò gần đây, hầu hết phụ nữ đều cho biết chu kỳ kinh nguyệt hằng tháng khiến họ cảm thấy thật sự mệt mỏi và khó chịu.
-
Dị tật ở cơ quan Sinh d*c nữ là những bất thường về cấu trúc của cơ quan Sinh d*c. Do thiếu hiểu biết hoặc do xấu hổ nên nhiều người không chịu đi khám khiến cho việc điều trị,
-
Người cao tuổi (NCT) thường nhìn vấn đề T*nh d*c như là một cách biểu lộ sự yêu mến, ngưỡng mộ và chung thủy, một cách thể hiện sự lãng mạn mới, một khẳng định của tình yêu, những cơ hội trải nghiệm trong cuộc sống.
-
Cuộc sống tinh thần ở người cao tuổi (NCT) có tính chất nền tảng, quyết định điều khiến nhịp độ lão hóa. Ngược lại, mục đích cơ bản của việc làm chậm sự lão hóa, kéo dài tuổi thọ là nhằm kéo dài thời gian lao động có ích và ước vọng sống lâu, khỏe mạnh.
-
Như một phép lạ kỳ diệu, bé trai 3 tuổi Adonis Ortiz ở Mỹ đã hồi phục mau chóng sau khi cấy ghép tới 5 cơ quan nội tạng.
-
Mô hình “Tuyến phố hiến máu” do Viện Huyết học và Truyền máu TW tổ chức đã đi vào hoạt động trên địa bàn TP. Hà Nội.