Con đường hoạt hóa bổ thể cổ điển (classical pathway)
Tin liên quan
- 10 công thức kết hợp màu sắc cổ điển giúp bạn có vẻ ngoài hoàn hảo (27/11/2020)
- Cô gái với sở thích kỳ lạ khi chỉ mặc trang phục cổ điển có từ thế kỷ 19 (23/11/2020)
- Hóa thành quý cô kiêu sa với những chiếc đầm mang vẻ đẹp cổ điển vĩnh cữu (06/11/2020)
- Lê Âu Ngân Anh khoe trọn vẻ thanh xuân ngọt ngào với style cổ điển (01/11/2020)
- Người đàn ông biến mọi thiết bị cơ khí cổ điển thành... nhện (13/11/2020)
Quá trình enzym và tấn công màng dẫn đến giết ch*t tế bào. Đơn vị nhận diện bổ thể hoạt động của con đường này có thể chia làm ba giai đoạn: nhận dạng, hoạt hóa là phức hợp C1.
Cố định C1 bởi immunoglobulin
Cố định C1 xảy ra khi tiểu đơn vị C1q gắn trực tiếp lên Ig. Hai tiểu đơn vị còn lại của C1 là Clr và Cls không kiên kết với Ig mà tham gia vào quá trình hoạt hóa cổ điển về sau. Phân tử kháng thể sau khi kết hợp với kháng nguyên tương ứng sẽ bị thay đổi cấu trúc không gian làm lộ các thụ thể đối với C1q. Tính chất kết hợp của bổ thể vào phức hợp kháng nguyên - kháng thể là cơ sở của phản ứng cố định bổ thể mà chúng ta thường hay sử dụng trong các xét nghiệm miễn dịch học.
Không phải tất cả các Ig có khả năng kết hợp với bổ thể. Ở người chỉ có IgG1, IgG3, và IgM là có khả năng này; ở chuột nhắt có IGg2 và IgM, và chuột lang có IgG2, .v.v. Cấu hình không gian của phức hợp kháng nguyên - kháng thể cũng có ảnh hưởng đến khả năng cố định bổ thể. Nhưng cụ thể thì chưa được biết rõ.
Một phân tử IgM pentamer kết hợp với kháng nguyên là có thể cố định bổ thể nhưng đối với IgG thì phải có phân tử IgG được gắn với kháng nguyên ở vị trí gần nhau mới có thể cố định được bổ thể. Để đạt được điều này trong thực nghiệm đối với hồng cầu cừu, người ta tính ra phải cần đến 600-800 phân tử IgG gắn lên hồng cầu. Do đó mà người ta thường dùng IgM hơn. Vị trí gắn của bổ thể vào Ig là lĩnh vực CH2 của IgG và CH4 của IgM. Sau khi C1q đã kết hợp với kháng thể, sự hoạt hóa sẽ đựơc truyền sang C1r và Cls. Cơ chế C1q hoạt hóa C1r vẫn còn chưa được biết. C1s sau khi được hoạt hóa C4 và C2 là hai thành phần tiếp theo của chuỗi phản ứng.
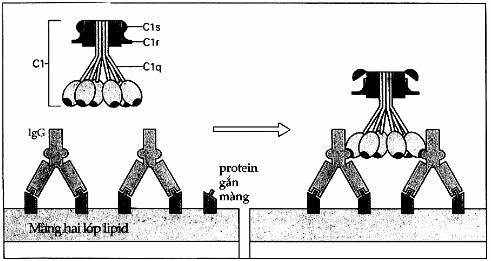
Hình. Cố định C1qrs.
Một cặp phân tử IgG liên kết với các phân tử protein kháng nguyên (protein gắn màng). Mảnh C1 cố định các lĩnh vực CH2 của IgG. Sự hoạt hóa C1r và C1s là do phân cắt nội bộ và do sự biến đổi của thành phần C1q tạo nên.
Cố định và hoạt hóa C4 và C2 bởi phức hợp C1qrs
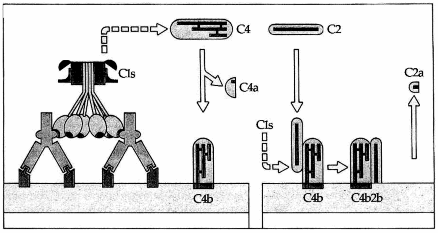
Hình. Sự hình thành C4b2b.
C1s cắt C4 tạo ra C4a và C4b. C4b gắn vào màng tế bào. C2 huyết thanh đến gắn vào C4b và bị C1s cắt để giải phóng C2a. C4b2b chính là enzym cắt C3 của con đường hoạt hóa cổ điển.
Phân tử C1s sau khi được hoạt hóa nó trở thành một enzym và cắt phân tử C4 thành hai mảnh: mảnh nhỏ là C4a và mảnh lớn C4b. Mảnh C4b có một vị trí hoạt động giúp nó gắn lên thụ thể dành cho nó trên bề mặt một số tế bào. Chúng ta cần lưu ý rằng mảnh C4b không nhất thiết chỉ nằm chung quanh nơi nó được hình thành mà nó thể vào máu tuần hoàn và được lưu động, tuy nhiên khi nó vào tuần hoàn thì trở nên bị bất hoạt. Phần C4b còn lại tại chäù (khoảng 10%) sẽ gắn lên tế bào.
C1s có tác dụng rất yếu lên C2 tự do, nhưng khi C2 tạo phức hợp với C4b thì tác dụng này mạnh hơn nhiều. Sự hấp thụ C2 lên C4b chỉ xảy ra trong điều kiện có mặt ion Mg . C2 sẽ bị cắt làm hai mảnh, mảnh nhỏ C2a sẽ bị mất vào môi trường chung quanh còn mảnh lớn C2b thì vẫn còn bám trên C4b tạo thành phức hợp C4b2b có tính enzym không bền vững, có thời gian nửa đời sống ngắn (5 phút) vì mảnh C2b dễ bị hủy hoại hoặc tách rời khỏi C4b (Hình 6.2).
Tác động của C4b2b lên C3
Phức hợp C4b2b hoạt hóa phân tử C3 bằng cách cắt phân tử này thành hai mảnh C3a và C3b. Mảnh C3a có trọng lượng phân tử thấp, được giải phóng vào trong dịch cơ thể và có hoạt tính của một hóa chất trung gian của phản viêm. Mảnh lớn của C3b có trọng lượng phân tử khoảng 175.000 kD và có khả năng dính lên bề mặt tế bào. Tuy nhiên, chỉ có tỷ lệ nhỏ C3b dính được vào màng tế bào ở vị trí sát với phức hợp enzym C4b2b để tạo phức hợp C4b2b3b có diện tác dụng nằm trên mảnh C3b. Các phân tử C3b bám vào màng tế bào ở các vị trí xa C4b2b thì chỉ có vai trò giúp cho việc kết dính miễn dịch và làm dễ hiện tượng thực bào nhờ vào đầu kia gắn lên thụ thể dành cho C3 ở các thực bào.
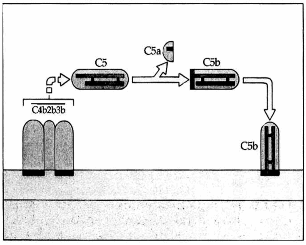
Hình. Cố định C5.
C4b2b3b tách mảnh C5a từ chuỗi α của C5 ra. Mảnh còn lại là C5b sau đó gắn vào màng tế vào.
Tác động của C3b lên C5
Phức hợp C4b2b3b mà diện tác dụng là C3b cắt phân tử C5 thành ra hai mảnh. Mảnh C5a là mảnh nhỏ cũng được giải phóng vào dịch cơ thể và có tác dụng như một chất trung gian hóa học của phản ứng viêm. Mảnh C5b sẽ được cố định lên màng tế bào và bắt đầu tạo phức hợp tấn công màng.
Nguồn: Internet.Chủ đề liên quan:
bổ thể classical pathway cổ điển con đường hoạt hóa bổ thể cổ điển hoạt hóa bổ thể cổ điểnTin liên quan
- “Nàng thơ” Lương Thuỳ Linh “hiện thực hoá những ý tưởng mộng mơ” qua ánh nhìn cổ điển (30/05/2020)
- 10 mẫu váy cổ điển mùa hè (08/06/2022)
- Hương Giang tựa như đóa hoa hồng nhung đen, đẹp mê hồn trong chiếc đầm cổ điển (19/10/2020)
- Lan Khuê, Mai Phương Thúy cho đến Selena Gomez đều hút hồn fan với đồ bơi cổ điển (29/05/2020)
- Ngôi nhà 30m² thiết kế cổ điển, hớp hồn những anh chàng độc thân (11/04/2020)
Tin cùng chuyên mục
Tin cùng nội dung
-
Các protein của hệ thống bổ thể tạo thành hai chuỗi enzym mà người ta gọi là con đường cổ điển và con đường không cổ điển để tạo nên hai cách phân cách hai phân tử C3 tức phân tử trung tâm của hệ thống bổ thể.
-
C8, C9 hai thành phần cuối cùng bị hoạt hoá sẽ tạo ra các lỗ thủng làm thay đổi tính thấm màng tế bào, làm tế bào trương to và ch*t.
-
Trong hệ thống 1, kháng nguyên được cho tác dụng với kháng thể. Nếu kháng nguyên và kháng thể phản ứng đặc hiệu thì tất cả lượng bổ thể kết hợp vào phức hợp kháng nguyên kháng thể
-
Nếu bổ thể hoạt hóa theo đường cổ điển như trong bệnh viêm thận lupus, định lượng nồng độ bổ thể huyết thanh C3, C4 và CH50 sẽ thấp
-
Bệnh nhân bị bệnh hồng cầu hình liềm dễ mắc phải tình trạng giảm chức năng con đường bổ thể thay thế
-
Bất kể được hoạt hóa theo con đường nào, sản phẩm cuối cùng được tạo ra sẽ là một phức chất protein có khả năng phá hủy màng tế bào của kháng nguyên.
-
Nếu như những năm trước xu hướng mũ có thiêng về snapback thì trong vòng 2 thái trở lại đây chính là thời điểm hot của mũ lưỡi trai cổ điển.
-
Hiện nay, phong cách cổ điển châu Âu rất được ưa chuộng, với các bức họa treo trong phòng ăn và phòng khách có thể giúp căn nhà bạn thêm sang trọng.
-
Trước khi các hãng mỹ phẩm ra đời, rất nhiều thứ đã được sử dụng để làm tăng vẻ đẹp cho phụ nữ. Dưới đây là vài mẹo làm đẹp các bạn nên xem.
-
Không có gì an toàn hơn việc chia sẻ với bạn tình bằng cách nghe nhạc khi quan hệ T*nh d*c của hai người đang vào thời điểm mật thiết nhất.